Part of our series of posts by final-year undergraduate students for their Research Comprehension module. Students write blogs inspired by guest lecturers in our Evolutionary Biology and Ecology seminar series in the School of Natural Sciences.
This week; views from Sarah Byrne and Sean Meehan on Tom Ezard’s seminar, Birth, death and macroevolutionary consequences.
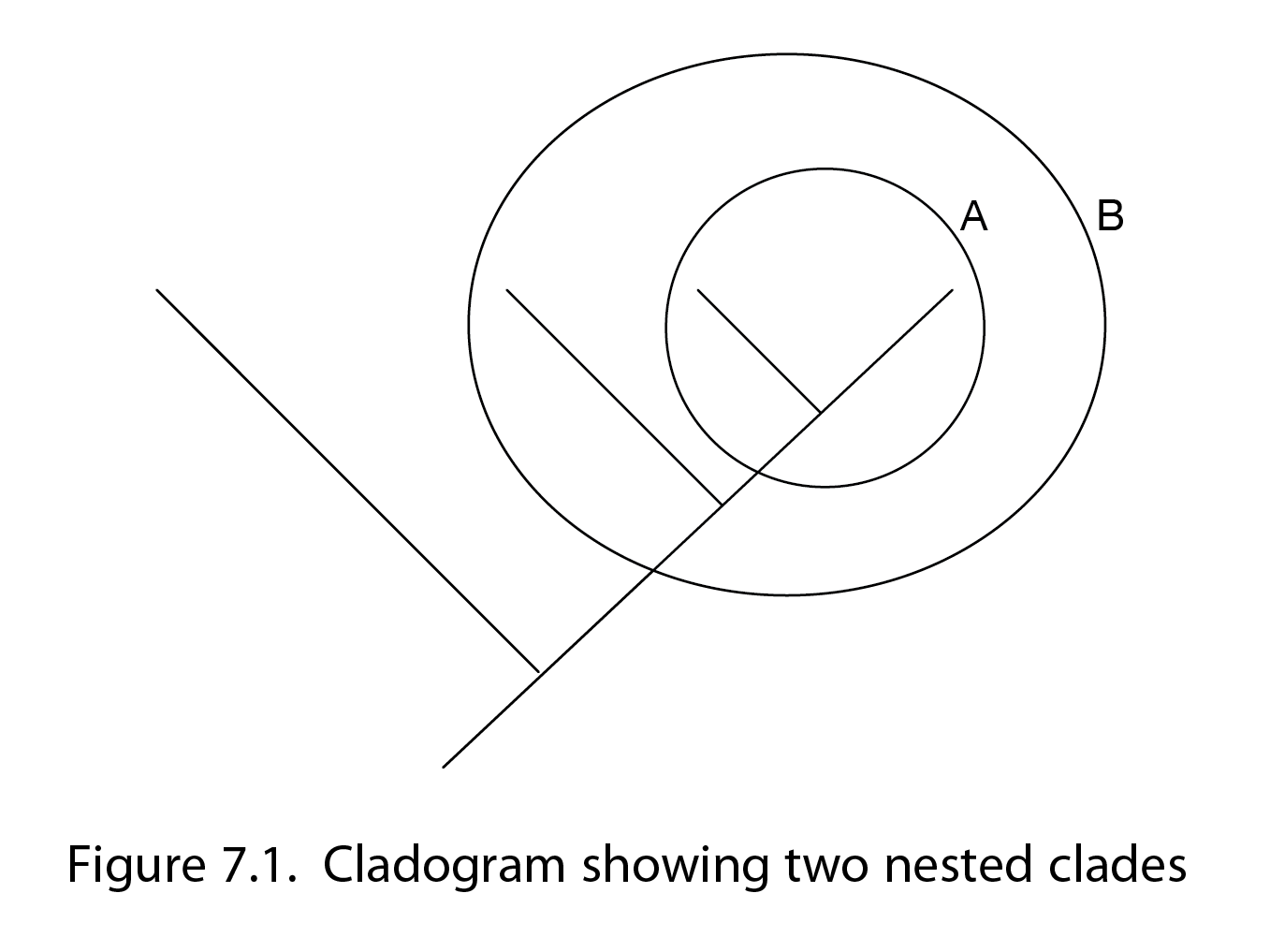
Splitting Hares – easier said than done?
In a recent talk given by Tom Ezard, a research fellow and evolutionary ecologist, the definition of a species was examined and challenged. While defining a species may seem a simple task for just about anybody and in particular a room full of people with a biology background, the actual definition can be harder to understand when thinking about fossil or species’ records and gaps across time. Ezard highlights that a dynamic approach is needed when discussing speciation and the definition of a species. Claiming that you shouldn’t define a species at one particular moment in time, he details that large gaps in the fossil record make it very difficult to have a fully complete picture about speciation events. In other words, making inferences about speciation events from a certain snapshot in time could overlook the dynamic process of change that occurs over time and give us inaccurate theories about the macroevolution of species.
Following on from the definition of a species, Ezard was interested in the fossil record and how it can give us information about the species record and also, more importantly, about diversity. He was interested in finding out where these gaps in the fossil record had occurred and what impacts they could possibly have. In graphs he provided, it was clear that there was a difference between data over time with more species surges found in recent data in comparison with the past, indicating the number of species has increased over time. However, it’s a little misleading because as time develops we learn more about how to indentify species of have better techniques to do so, it is therefore unclear as to whether or not there has been a big increase in species.
To better explain some complicated parts of the speciation theory, Ezard used a baseball analogy which I was thankful for, showing a picture of various baseballs over time. Ezard explained how techniques improve over time and how the original was very different to the new and modern ball. All of the baseballs of various different ages, textures and shapes remained part of one game (or one species) and that there was no split into a new game (or new species). He stressed that this continuation was very important in understanding macroevolution and when identifying species, that it was vital to look at gaps in the lineage. This brings us back to the fact that the fossil record needs to be examined further and the question of what is meant by a species may need to be redefined. Ezards definition of a species as ‘a single line of descent, a sequence of populations evolving separately from others seems closer to the real definition than previously thought.
Speciation was also a key factor of Ezard’s talk and he was interested in identifying budding speciation events while still being able to identify their ancestors. Two main types of speciation and evolution were discussed in the talk, one type; anagenesis refers to a change along a branch of a phylogeny or the evolution of a gradual change within a species over time. This theory was backed by Darwin and eventually leads to a speciation event. In contrast, cladogenesis, where a population stays stable until a big speciation event happens suddenly and then a splitting occurs between species that ensures they can then not reproduce with each other.
The split can be caused by either biotic or abiotic factors with disagreements regularly occurring between geologists and modern evolutionary biologists over whether the biotic factors (such as competition) or the abiotic factors (such as climate) are the main key drivers affecting species ecology and diversification. So, what is the main driver affecting species ecology and in turn speciation and diversification? Ezard was interested in finding this out.
Using observational studies, algorithmic processes and a multivariate complex approach, Ezard was able to account for ecological differences between species. Lotka’s equation gave an estimate of birth and death models that detailed speciation probability and extinction risk. Species respond differently to global drivers of change and these differences have macroevolutionary consequences. The Red Queen Hypothesis mentioned above, a biotic factor that describes how predator and prey are continually adapting to out-do each other affects species much more so than climate does, and in comparison, climate, an abiotic factor has much more of an effect on extinction.
So, it seems that a combination of both factors are important although they affect both speciation and extinction at different rates. Ezard indicated that, in order to understand diversity, it was first necessary to understand the biotic factors that impact the split and to then devise a model to draw these two areas together. Ezard’s enthusiastic and engaging approach clearly showed his passion for the subject and the interesting topic left me with a lot to think about it.
Author: Sarah Byrne
——————————————————————-
Lumpers and Splitters: Apparently they’re not varieties of potato
What is a species? This question seems so fundamental to biology that surely the experts have answered it by now, right? Wrong. Defining a species is a difficult thing, and each new definition seems to come up short in certain criteria. For example Ernst Mayr’s widely used definition of a species: “groups of actually or potentially interbreeding natural populations, which are reproductively isolated from other such groups” completely disregards species which reproduce asexually. For this reason I like Simpson’s evolutionary concept for defining a species and this is precisely what Tom Ezard uses for his work on macroevolutionary dynamics. This concept holds that each species represents a single line of descent; it begins with a speciation event and terminates with extinction. Ezard used the evolution of the baseball to demonstrate this concept. Although the modern baseball is considerably different from its original ancestor, it is still a baseball and there have been no ‘speciation’ events or splits in the lineage to form a new type of ball.
It was Darwin who first coined the term ‘lumpers and splitters’. Lumpers are those biologists who tend to ‘lump’ many ‘species’ in together as one. The splitters are those biologists who like to make as many ‘species’ as possible. In his 1945 work ‘The Principles of Classification and a Classification of Mammals’ George G. Simpson notes rather sardonically: “splitters make very small units – their critics say that if they can tell two animals apart, they place them in different genera … and if they cannot tell them apart, they place them in different species. … Lumpers make large units – their critics say that if a carnivore is neither a dog nor a bear, they call it a cat.” So we can see that this problem is an old one, and that Simpson’s evolutionary concept is very useful for defining species in macroevolutionary studies.
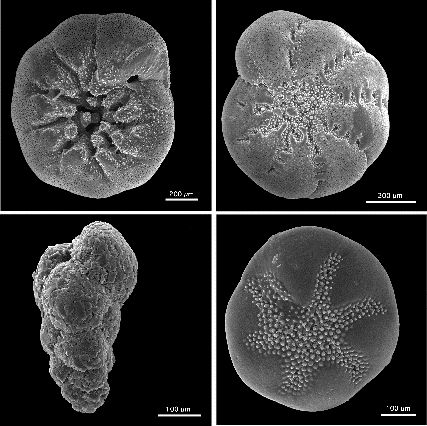
In order to study macroevolutionary dynamics one needs a fairly detailed picture of a clade’s development, and not many organisms provide a suitable fossil record for a detailed study. Fortunately Ezard and his team found the perfect organisms for this purpose; the Foraminifera. These creatures are marine dwelling amoeboid protists. When they die they sink to the bottom and leave behind their calcium shells or tests. They are deposited and preserved on the sea floor and in the right conditions over time can form stratified layers of fossils which give a very complete picture of their evolution over time. Also,the stable isotope ratios of oxygen in the shells can be used to reconstruct palaeo-climatic conditions. These attributes make them incredibly useful in the study of macroevolutionary dynamics.
So, what are the driving forces of speciation? Is there one factor which influences this process above all the others? This is what Ezard and his team set out to investigate. The foraminifera had an interesting story to tell. It was found that incipient species diversify the fastest. This was found to be primarily due to biotic factors or ‘Red Queen’ factors. As a clade grows older it was found that diversification slows due to diversity dependence. However, it was found that extinction is primarily influenced by climatic or Court Jester factors. These findings are important in order to grasp a general understanding of macroevolutionary dynamics. It means that impacts of diversity and climatic fluctuations are not felt uniformly across a phylogeny. More simply put, it means that the extent of the effect of biotic and abiotic factors on a clade depend on how old it is.
In summary, what Ezard and his team found was that there is no dominant macroevolutionary force, but that, a combination of biotic and abiotic variables drive speciation and extinction. They also found that species’ ecologies are important driving forces in these processes.
Author: Sean Meehan
Image Source: Wikicommons